First, we wish to
simulate the complete iFFL process and explore the feasibility of using the CRISPR-CasF system as a
repressor in the iFFL process.
However, since we
still do not know much about CasF, we decided to model CasF using dCas9, the most commonly used
CRISPRi system. Existing studies have measured the metrics of dCas9, which will facilitate our
modeling.
The complete iFFL
regulatory process concerning target gene expression is shown below.
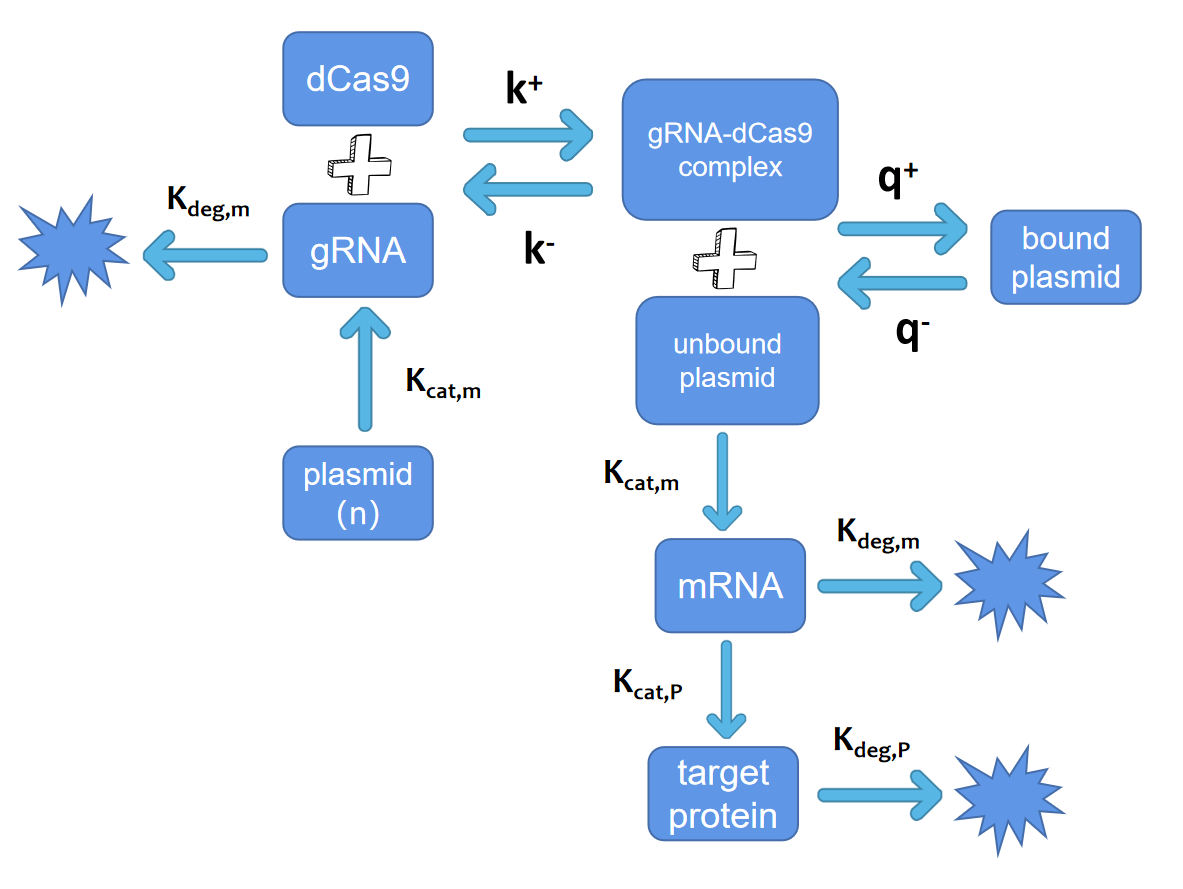
Fig. 3.
Elucidation of the primary reactions in iFFL system
The above process is
expressed in the assumption that we use the following ordinary differential equation:
$$
\frac{d}{dt}\left[ g \right] =\left[ n \right] u+K^-\left[ c \right] -K^+\left[ d \right] \left[ g
\right] -\frac{K_{d,m}\left[ g \right]}{K_{M,m}+\left[ g \right]}\tag{6}
$$
$$
\frac{d}{dt}\left[ c \right] =K^+\left[ d \right] \left[ g \right] -K^-\left[ c \right] +q^-\left[ C
\right] -q^+\left[ c \right] \left[ D \right] \approx K^+\left[ d \right] \left[ g \right] -K^-\left[ c
\right] \tag{7}
$$
$$
\frac{d}{dt}\left[ C \right] =q^+\left[ c \right] \left[ D \right] -q^-\left[ C \right] \tag{8}
$$
$$
\begin{aligned}
\frac{d}{dt}\left[ m \right] &=\frac{K_{cat,m}\left[ D \right] \left[ E_{70} \right]}{K_{M,70}+\left[
E_{70} \right]}-\frac{K_{d,m}\left[ m \right]}{K_{M,m}+\left[ m \right]}\\
&\approx K_{cat,m}\left[ D \right] -K_{deg,m}\left[ m \right] \left( K_{M,70}\ll \left[ E_{70} \right]
,\mathrm{ }K_{M,m}\gg \left[ m \right] \right)\\
\end{aligned}\tag{9}
$$
$$
\frac{d}{dt}\left[ Y \right] =\frac{K_{cat,P}\left[ m \right] \left[ R_0 \right]}{K_{M,R}+\left[ R_0
\right]}-K_{deg,p}\left[ Y \right] \approx K_{cat,P}\left[ m \right] -K_{deg,p}\left[ Y \right] \left(
K_{M,R}\ll \left[ R_0 \right] \right) \tag{10}
$$
Where $
\left[ n \right]
$ is the plasmid concentration, $u$ is the gRNA transcription rate, $
\left[ d \right]
$ is the dCas9 concentration, $
\left[ g \right]
$ is the gRNA concentration, $
\left[ c \right]
$ is the gRNA and dCas9 complex concentration, $
\left[ C \right]
$ is the concentration of the Cas protein complex bound to the target sequence, $
\left[ D \right]
$ is the unbound target gene concentration, $
\left[ m \right]
$ is the mRNA concentration transcribed by target gene, $
\left[ Y \right]
$ is the concentration of the target protein, $
K_{d,m}=K\left[ R_{nase} \right]
$, $
k^+
$ and $
k^-
$ are decomposition and binding rate of gRNA and Cas protein, $
q^+
$ and $
q^-
$ are are decomposition and binding rate of gRNA-Cas complex and target sequence.
According to our
definition:
$$
\left[ n \right] =\left[ D \right] +\left[ C \right] \tag{11}
$$
$$
\left[ D \right] =\left[ P \right] \tag{12}
$$
When the cell growth
is stable, the gRNA concentration, gRNA-dCas9 complex concentration, gRNA-dCas9 complex and target
gene binder concentration, unbound target gene concentration, mRNA concentration, and target protein
concentration all reach a steady-state. Solving equations $
(6)
$ and $
(7)
$:
$$
\left[ c \right] =\frac{\left[ d \right] \left[ g \right]}{K}\tag{13}
$$
$$
\left[ C \right] =\frac{\left[ d \right] \left[ g \right] \left[ D \right]}{KQ}\tag{14}
$$
Where $
K=\frac{k^+}{k^-},Q=\frac{q^+}{q^-}
$.
Relating to equation
$
(10)
$, it follows that:
$$
\left[ D \right] =\frac{\left[ n \right]}{1+\frac{\left[ d \right] \left[ g \right]}{KQ}}\tag{15}
$$
Taking equation
$(11)$ back to equation $(5)$:
$$
\frac{d}{dt}\left[ g \right] =\left[ n \right] u-\frac{K_{d,m}\left[ g \right]}{K_{M,m}+\left[ g
\right]}\tag{16}
$$
Solving this
equation, we get:
$$
\left[ g \right] _{SS}=\frac{\left[ n \right] u}{K_{deg,m}}\tag{17}
$$
Substituting equation
$(15)$ into equation $(13)$:
$$
\left[ D \right] _{SS}=\frac{\left[ n \right]}{1+\frac{\left[ d \right] \left[ n \right]
u}{KQK_{deg,m}}}=\frac{1}{\frac{1}{\left[ n \right]}+\frac{\left[ d \right] u}{KQK_{deg,m}}}\tag{18}
$$
Substituting equation
$(16)$ into equation $(13)$, the final stable concentration of the target protein:
$$
\left[ Y \right] _{SS}=\frac{K_{cat,m}}{K_{deg,m}}\cdot \frac{K_{cat,P}}{K_{deg,P}}\left[ D \right]
=\frac{K_{cat,m}}{K_{deg,m}}\cdot \frac{K_{cat,P}}{K_{deg,P}}\cdot \frac{1}{\frac{1}{\left[ n
\right]}+\frac{\left[ d \right] u}{KQK_{deg,m}}}\,\tag{19}
$$