Team:NUS Singapore/Description
Project Description
Overview

Our PRYSM biomanufacturing system takes advantage of optogenetics to interface hardware and biological systems for the precise and convenient control of 3 key functions in our engineered Saccharomyces cerevisiae.
- The application of red light induces the yeast to produce and secrete our biopesticide of choice: Human beta defensin.
- The application of blue light induces the yeast to flocculate, causing them to aggregate and sediment to allow for convenient extraction of our peptide.
- Concurrent use of red and blue light activates a nuclease based kill switch, allow easy sterilization of extracted media and adds an insulating layer of biosafety.
Design Principles
Our project was designed with the aim of satisfying the following technical goals, click on the icons to find out more:






Design Choices
Human beta defensin
Defensins belong to the category of antimicrobial peptides (AMPs), which have a non-enzymatic inhibitory effect on a broad spectrum of microorganisms [9]. Despite the fact that this class of peptides has a wide range of structures, their antibacterial properties are all generated from comparable mechanisms. These peptides interact electrostatically with negatively charged microbial cell membrane components (e.g., phospholipids) due to their positive charge, increasing membrane permeability and eventually leading to cell death.
We chose to use Human Beta Defensin 2 (HBD2) as our primary candidate due to several reasons. Firstly, functional recombinant HBD2 has been demonstrated expressed using S.cerevisiae. However, to date, no practical application of recombinant HBD2 has been attempted [3]. Furthermore, most works regarding AMPs and HBD2 have been focused on the therapeutic field. We decided that it would be interesting to test the broad-spectrum antimicrobial activity demonstrated in medical research in the agricultural context. Lastly, HBD2 is native to the human body, rendering it extremely human-safe.
Secondly, while performing our integrated human practices, we consulted Vaciome, a Singaporean synbio startup specializing in AMP discovery and production. Through our discussions, we discovered that they had an HB2 construct for Pichia pastoris that could easily be ported for S.cerevisiae. Upon our request, they graciously donated a sample of their HBD2 construct for our iGEM project, which helped us save resources that would have been used on synthesis.
Saccharomyces cerevisiae
Description: Saccharomycies cerevisiae is most commonly known as baker’s yeast! It is found throughout history and society, from bread baking to some of the most successful instances of industrial biomanufacturing.
We decided S.cerevisiae was an ideal chassis for the production of human beta defensin. As a model GRAS organism[1] that is heavily used in the bioproduction and food industry, it was undeniably the safest option for agricultural applications.
S.cerevisiae is also well characterized in protein production and genetic editing, with previous work demonstrating the successful production of human beta defensin using this chasis[2,3]. This meant that we could reliably produce high yields of human beta defensin.
Flocculation

Flocculation refers to a phenomenon most commonly observed in brewer’s yeast. After fermentation, yeast will aggregate and sink to the bottom of the barrel[4]. It is induced by the production and secretion of the protein Flo1, which is normally inactive in lab and industrial strain yeasts. The Flo1 protein on the surface of yeast bind to mannose residues in the cell wall of other yeasts, resulting in aggregation.
We realized that using this in conjunction with a secreted protein would allow us to separate out one of the most troublesome contaminants in the products of biomanufacturing: the cells themselves. This avoids incurring the costs of expensive chemicals or machinery needed for production, making it far more cost effective than conventional solid-liquid separation methods such as filtration, centrifugation or sedimentation.
In addition, this would limit the amount of genetically modified organisms that could end up in our product, adding an additional level of biosafety.

Human beta defensin can be cytotoxic to fungi[5]. Thus if constitutively expressed, may negatively impact the growth of the yeast and the overall biomass generated, decreasing yield in the long term. Flo1 has to be processed through the secretory pathways before inducing flocculation [6], possibly competing with the secretion of HBD.
Therefore, it is of paramount importance to have precise control over when we want to induce our production of these two genes!
Optogenetics
Optogenetics is the use of light to control gene expression, usually through the manipulation of proteins that alter their binding in the presence of a specific wavelength of light. Using optogenetics, different coloured lights can be used to turn specific genetic circuits on and off.
In order to balance the dual functionalities of beta defensin production and flocculation alonside cell growth, we aimed to implement a dual channel optogenetic system, such that red light activates the expression of human beta defensin, and blue light activates the expression of the Flo1 gene.
Why light?
Light is an attractive alternative to conventional chemical based inducers for several reasons
- It is much cheaper
- Administration of light can be automated
- Less cytotoxicity/safety hazards
- Minimal impact on cell metabolism and thus will not hinder cell growth or production
Red Light
The red light channel was designed based on the PhiRex system published by Potsdam University[7]. This system revolves around a split protein framework:
- The first protein is a fusion between a SynTALE DNA binding domain and a PhyB domain from the plant Arabidopsis thaliana
- The second is a fusion between the PIF3 domain and a VP64 activation domain
- The synthetic promoter consists of a truncated CYC1 promoter, taken from the yeast native genome, with it’s upstream activation sequence replaced with the DNA site targeted by the SynTALE DNA binding domain
The SynTALE DNA binding domain will bind upstream of the truncated CYC1 promoter, bringing with it the PhyB domain. In the presence of red light, unbound PIF3 VP64 fusion proteins form a complex with PhyB, bringing the activation domains into proximity to the core promoter, resulting in downstream transcription.
In our system, red light will be used to control the production of human beta defensin, and will only be switched on after the cell culture has grown to a significant biomass to offset the detrimental effects it may have on cell growth. This way we can obtain the maximum number of cells before beginning production of human beta defensin.

Blue Light
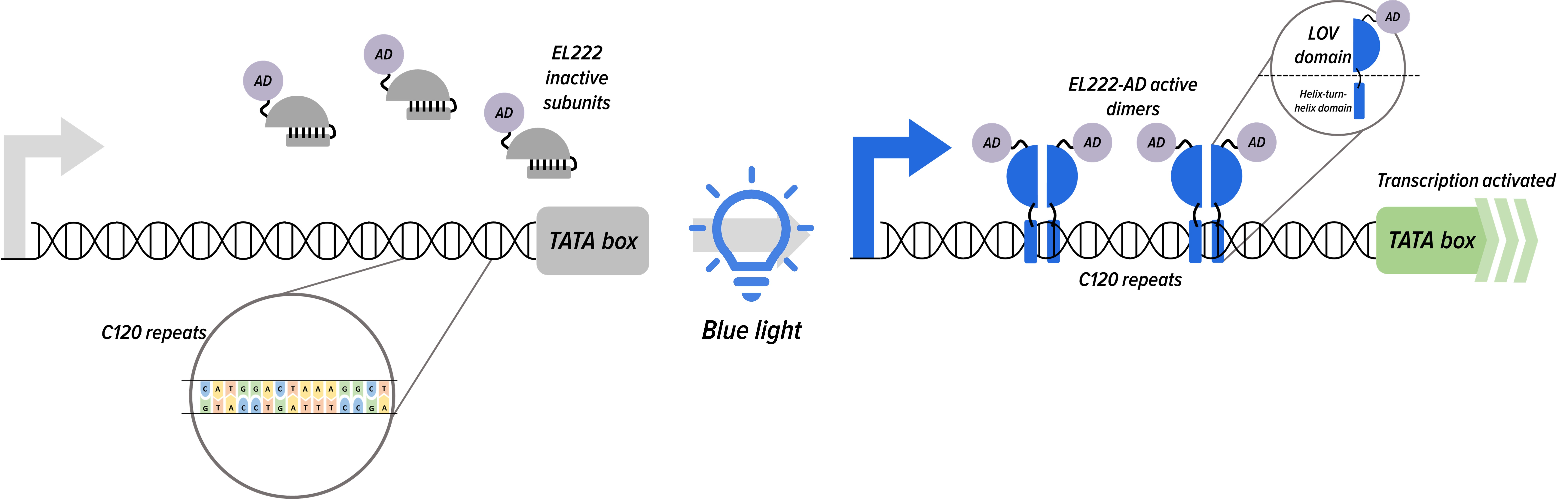
The blue light channel was designed based on the EL222 system for yeast published by Benzinger and Khammesh[8]. It consists of an EL222 protein fused to an NLS and a VP16 activation domain. In darkness, the EL222 proteins remain as a single subunit, but upon blue light exposure, EL222 subunits change their conformation, which exposes both a LOV domain and a helix turn helix domain, allowing them to form homodimers which bind to a target C120 sequence, bringing the activation domain into close proximity and activating downstream transcription.
In our system, blue light will be used to control the expression of flocculation such that the yeast will only divert resources and the secretory pathway towards the Flo1 protein after human beta defensin has been sufficiently produced.
Kill Switch
Biosafety is a key concern in synthetic biology, genetically modified organisms should have minimal opportunity to escape into the wild and in the event that they do, they should be modified to have severely impaired survivability.
Other than the organism itself escaping, it is also crucial that the modified genetic material does not escape into the environment for other organisms to use. To combine both of these concepts, we decided to implement an endonuclease based kill switch inspired by Team Sau Paulo, Brazil in 2017, that would simultaneously destroy our modified DNA as well as induce cell death.
Workflow of Reactor

References
- Bernardi, B., & Wendland, J. (2020). Homologous Recombination: A GRAS Yeast Genome Editing Tool. Fermentation, 6(2), 57. MDPI AG. Retrieved from http://dx.doi.org/10.3390/fermentation6020057
- Cipáková I, Hostinová E. Production of the human-beta-defensin using Saccharomyces cerevisiae as a host. Protein Pept Lett. 2005 Aug;12(6):551-4. doi: 10.2174/0929866054395761. PMID: 16101394.
- Møller, T.S.B., Hay, J., Saxton, M.J. et al. Human β-defensin-2 production from S. cerevisiae using the repressible MET17 promoter. Microb Cell Fact 16, 11 (2017). https://doi.org/10.1186/s12934-017-0627-7
- Soares, E. (2011), Flocculation in Saccharomyces cerevisiae: a review. Journal of Applied Microbiology, 110: 1-18. https://doi.org/10.1111/j.1365-2672.2010.04897.x
- Aerts, A.M., Thevissen, K., Bresseleers, S.M. et al. Arabidopsis thaliana plants expressing human beta-defensin-2 are more resistant to fungal attack: functional homology between plant and human defensins. Plant Cell Rep 26, 1391–1398 (2007). https://doi.org/10.1007/s00299-007-0329-4
- Bony M, Thines-Sempoux D, Barre P, Blondin B. Localization and cell surface anchoring of the Saccharomyces cerevisiae flocculation protein Flo1p. J Bacteriol. 1997 Aug;179(15):4929-36. doi: 10.1128/jb.179.15.4929-4936.1997. PMID: 9244284; PMCID: PMC179343.
- Hochrein, L., Machens, F., Messerschmidt, K., & Mueller-Roeber, B. (2017). PhiReX: a programmable and red light-regulated protein expression switch for yeast. Nucleic acids research, 45(15), 9193–9205. https://doi.org/10.1093/nar/gkx610
- Benzinger, D., Khammash, M. Pulsatile inputs achieve tunable attenuation of gene expression variability and graded multi-gene regulation. Nat Commun 9, 3521 (2018). https://doi.org/10.1038/s41467-018-05882-2
- Park, M. S., Kim, J. I., Lee, I., Park, S., Bae, J.-Y., & Park, M.-S. (2018). Towards the application of human defensins as antivirals. Biomolecules & Therapeutics, 26(3), 242–254. https://doi.org/10.4062/biomolther.2017.172
