Team:City of London UK/Engineering
Engineering
In order to create a working test for Preeclampsia, we designed multiple generations of Toehold switches and an isothermal amplification strategy in addition to hardware including a luminometer, magnetic nanoparticle extraction strategy and reaction chamber. However, this section will focus on the engineering success of our toehold switch and amplification probe design.
While we had access to labs at some points during our project, this came late on in our project, so much of our testing was done in Silico. On this page, we will outline two design cycles, the first using in Silico testing using our software tool, and the second using wet lab characterisation.
The First Cycle
Despite its name, this first cycle starts with our second generation (gen2) toehold switches, after we tested our gen1 switches using Nupack and then, after talking to Dr Alex Green and Dr Bae Wooli, a researchers in the field, we decided to create simpler, more specific ‘gen2’ switches. You can read more about this in our Design page.
Goal

Our research identified two miRNAs upregulated roughly 4-fold in patients with Preeclampsia at 10 weeks - miR-210-3p and miR-517-5p. So, the aim of the switches was to be able to discriminate between this percentage difference in concentration.
Design

Our gen2 toehold switches work such that when few microRNAs are present, the ribosome binding site of an mRNA coding for luciferase is folded up, so the ribosome cannot unzip the long stem and move down the strand to the start codon to initiate translation.
However, when a microRNA binds to the trigger site, the switch unfolds, decreasing the length of the stem, and exposing the start codon, so translation of leuciferase - a reporter protein - can begin.
We used the software package NUPACK to help design our gen2 switches in-silico. NUPACK is able to predict the interactions of the secondary structure of one or more nucleic acid strands. And so, we used the programme to model and perfect our switches.
At first, we designed our switches base by base, checking the minimum free energy (MFE) structure, using NUPACK, to ensure that our switch had a strong hairpin structure in the ‘off’ state and was properly unfolded in its ‘on’ state.
We were successful in designing multiple switches that had MFE structures fulfilling this criteria, but multiple bases had low probabilities of being in their respective positions.
Therefore, we used the NUPACK API to generate a Python program to test a randomly generated list of one-hundred-thousand linker regions (which did not contain any stop or start codons) and simulated a thousand probabilistic secondary structure samples for each, showing which switch had the highest probability of unfolding in the presence of our miRNA triggers, but not in their absence.
(To read more about specific design considerations, see the Design page).
The gen2 switch for miR-517-5p

The gen2 switch for miR-210-3p
Build
After this design, we ended up with two toehold switches (seen above) which we decided to characterise using our software tool, ToeholdTools. The building in this design cycle was for the software tool itself, rather than the switches, as we were testing them at first in Silico.
Test
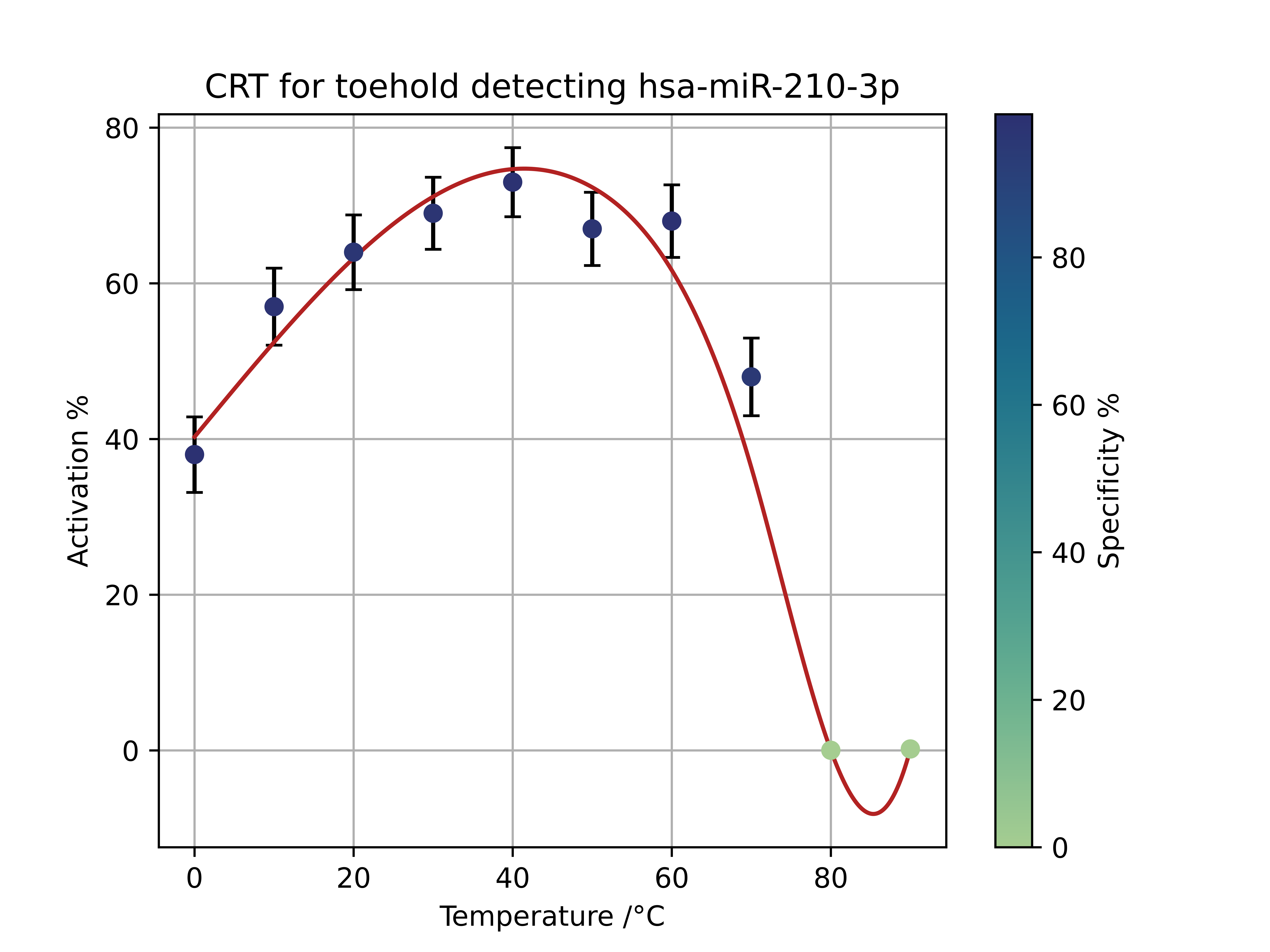

The software tool utilises Nupack’s API to test to see if the switches will unfold, or ‘activate’ in the presence of our target miRNA, but not in the presence of other miRNAs in the human miRNA database (which we downloaded from miRBase). The ideal switch is one that has high activation for our target miRNAs but not for other miRNAs. We found that both our switches performed well in silico - with high activation and relatively high specificity for the miR-210-3p switch, but less specificity for the miR-517-5p.
Learn
Based on these results, we decided we should design a protocol to characterise these switches in the lab, to see how they would unfold, thereby inducing translation of a luciferase CDS, emitting luminescence when luciferin is added, in the presence of our target miRNAs at a concentration, and a quarter of that concentration (to mimic the ratio in human serum between those with and without the condition) as well as in the presence of similar miRNAs, or ‘homologs’, that our software tool had identified as being able to somewhat unfold our switch.
Furthermore, this made us realise that in order to create a specific test, it would be better to be able to detect multiple miRNAs with the same switch, such that even if another miRNA homolog was over-expressed in the serum of a patient without the condition, it wouldn’t give a positive test unless all target miRNAs are expressed. To show a proof of concept for this idea, we got to work designing a novel AND-gate switch. This fed into our next design cycle.
The Second Cycle
Design
Our third generation (gen3) toehold switch uses AND-gate logic to detect the presence of our two target miRNAs in one switch.
This switch employs an anti-miRNA with three regions; two hybridization domains that are complementary to each of our trigger RNAs and the anti-miRNA’s own trigger site. This 29 nucleotide long strand has a 13 nucleotide long binding site for the miRNA 210-3p and an 11 nucleotide long binding site for miRNA 517-5p. This complex of the 3 RNA strands is necessary to bind to the toehold switch’s trigger binding site. We had to ensure that each of these trigger binding sites was >10 nucleotides on the advice of Dr Bae Wooli.


For the AND gate, we were unable to use the NUPACK API as the minimum free energy structure would show each strand binding, independent of the position of the other strands. Therefore, we designed the anti-miRNA and the toehold switch base by base. To ensure that the switch would unfold, we had to ensure that the difference in MFE structure between the ‘off’ and ‘on’ state was such that unfolding was energetically favourable when the anti-miRNA-miRNA complex had formed. In order to help us achieve this energetically favourable state we replaced an adequate amount of C-G bonds with U-G bonds to raise the free energy of the MFE structure of the OFF state.
Build
We ordered four gBlocks from IDT. The first three contained each of our two gen2 switches and also our gen3 switch. All three were downstream of a T7 promoter and upstream of a luciferase coding region. The final gBlock was for the anti-miRNA for the gen3 switch, which was also under the control of a T7 promoter. All these were linear DNA with 250 randomly generated base pairs upstream and downstream of the insert. This was on the advice of the supplier of the TXTL cell-free ktis we were using in our characterisation.
Test
To find out more about the performance of our gen2 switches, go to our Measurement and Results page. But, as they are not part of this design cycle, we don’t anaylse their results here.
The main purpose of testing the AND-Gate Switch was to see whether it could discriminate between a high concentration of one miRNA, and a low concentration of another, and a high concentration of both. This was in case a patient had increased levels of one miRNA and not the other, which would not be indicative of Preeclampsia. This would also show that our concept could potentially be scaled up, in order to create a framework for multiple miRNAs to be tested for, increasing the specificity of tests.


The graphs show how, compared to the negative control, which shows leaky expression of the toehold switch, the luminescence output of the sample with 9M concentration of both miRNA triggers is 92% higher. Interestingly, the percentage increase in luminescence of the miR-210-3p at 9M and miR-517-5p at 2.25M is twice more than twice as high as the miRNAs at flipped concentrations - 28% compared to 12%. This could be because the miR-210-3p binding site is further downstream than the miR-517-5p binding site, so miR-210-3p is able to bind and partially unfold the switch in the absence of miR-517-5p, increasing the rate of translation. However, as the miR-517-5p binding site is further upstream, it is completely bound up in hydrogen bonds, so it is not accessible to the miRNA unless it binds as the switch is being transcribed.
Learn
We concluded that the gen3 toehold switches do cause a noticeable increase in translation when both miRNAs are at a high concentration (9M), compared to when one of both of them is at a low concentration (2.25M), which gives strong evidence that our AND-gate design works. However, we realised that, since the 2.25M concentrations only caused a small amount more translation of luciferase than the negative control (due to leaky expression), if the concentrations in human serum were lower than that, we would not be able to discriminate between patients with or without preeclampsia, despite there being a 4-fold increase in patients with the condition. Thus, we decided we needed to redesign our test as a whole by incorporating a method of amplifying miRNAs before adding toehold switches to the example.
Design
We wanted to find a solution that would allow amplification to take place isothermally, in a single tube, increasing the accessibility of our tests, especially in countries with less access to expensive equipment such as thermocyclers.
Further research introduced us to miRPA which could convert miRNAs into double stranded DNAs with the miRNA encoded in part of one strand in DNA. In order to be left with just the strand with the miRNA encoded in it, we decided our kit would use another method on top of this - asymmetric RPA, which works the same as RPA, but with an excess of the forward primer, such that an excess of the desired strand is polymerised. To learn more about our design considerations, and the python tool we created to help us design probes, visit the Isothermal Amplification section of our Design page.