MlrA, or microcystinase, cleaves the peptide backbone of MC-LR cyanotoxins produced by microcystis aeruginosa cyanobacteria. This crucial step results in the peptide losing the cyclic structure, and therefore the majority of its toxicity. Further degradation continues in an enzymatic pathway by interactions with other peptidolytic enzymes from the microcystinase gene cluster. This part originally comes from the Aalto-Helsinki 2016 iGEM team (Part BBa_K1907001), but was originally codon-optimized for yeast.
Maturation Protein A is required in order for the capsid of the MS2 phage, and subsequently MS2-derived phage-like particles (PLP), to bind to bacterial pili. It is released from the capsid upon binding, and enters the cell along with the cargo to be delivered by the MS2 capsid.
Maturation Protein A is required in order for the capsid of the MS2 phage, and subsequently MS2-derived phage-like particles (PLP), to bind to bacterial pili. It is released from the capsid upon binding, and enters the cell along with the cargo to be delivered by the MS2 capsid. This maturation protein A coding region includes a histidine tag on the N-terminus, to allow for purification as a monomer for capsid assembly in vitro.
This part is a modified version of BBa_K3738000, to which an N-terminal histidine tag has been added. As maturation protein A has not been well studied for use in phage-like particles, we have designed various versions of the part for testing purposes.
Maturation Protein A is required in order for the capsid of the MS2 phage, and subsequently MS2-derived phage-like particles (PLP), to bind to bacterial pili. It is released from the capsid upon binding, and enters the cell along with the cargo to be delivered by the MS2 capsid. This maturation protein A coding region includes a histidine tag on the C-terminus, to allow for purification as a monomer for capsid assembly in vitro.
This part is a modified version of BBa_K3738000, to which an C-terminal histidine tag has been added. As maturation protein A has not been well studied for use in phage-like particles, we have designed various versions of the part for testing purposes.
Maturation Protein A is required in order for the capsid of the MS2 phage, and subsequently MS2-derived phage-like particles (PLP), to bind to bacterial pili. It is released from the capsid upon binding, and enters the cell along with the cargo to be delivered by the MS2 capsid. This maturation protein A coding region includes a histidine tag on the N-terminus, to allow for purification as a monomer for capsid assembly in vitro. There is also a TEV cleavage site between the coding region and the histidine tag, in order to allow for histidine tag removal following purification if desired.
This part is a modified version of BBa_K3738000, to which an N-terminal histidine tag and TEV cleavage site have been added. As maturation protein A has not been well studied for use in phage-like particles, we have designed various versions of the part for testing purposes.
Maturation Protein A is required in order for the capsid of the MS2 phage, and subsequently MS2-derived phage-like particles (PLP), to bind to bacterial pili. It is released from the capsid upon binding, and enters the cell along with the cargo to be delivered by the MS2 capsid. This maturation protein A coding region includes a histidine tag on the C-terminus, to allow for purification as a monomer for capsid assembly in vitro. There is also a TEV cleavage site between the coding region and the histidine tag, in order to allow for histidine tag removal following purification if desired.
This part is a modified version of BBa_K3738000, to which an C-terminal histidine tag and TEV cleavage site have been added. As maturation protein A has not been well studied for use in phage-like particles, we have designed various versions of the part for testing purposes.
The MS2 capsid protein comprises most of the capsid of the MS2 phage, as well as MS2 phage-like particles (PLPs). A single MS2 capsid requires 178 copies of the capsid protein, as well as one copy of maturation protein A (enter part number here), which is critical in allowing the capsid to bind to the cell and deliver its cargo. This part is a coding region comprised of 2 copies of the MS2 capsid protein, joined by a histidine tag which is flanked by linker sequences (de Martin Garrido et al., 2020). This part has been codon optimized for use in E.coli.
The MS2 capsid protein comprises most of the capsid of the MS2 phage, as well as MS2 phage-like particles (PLPs). A single MS2 capsid requires 178 copies of the capsid protein, as one copy of maturation protein A (BBa_K3738000), which is critical in allowing the capsid to bind to the cell and deliver its cargo.
The design of this part was based of part BBa_K2683012, which is an MS2 capsid protein fused to a polyarginine tag cell-penetrating peptide on the N-terminus. This part has been modified from the original so as to have the polyarginine tag replaced with a histidine tag, to allow for purification as a monomer for capsid assembly in vitro. This part has been codon-optimized for use in E.coli.
The MS2 capsid protein comprises most of the capsid of the MS2 phage, as well as MS2 phage-like particles (PLPs). A single MS2 capsid requires 178 copies of the capsid protein, as one copy of maturation protein A (BBa_K3738000), which is critical in allowing the capsid to bind to the cell and deliver its cargo. The design of this part was based of part BBa_K2683012, which is an MS2 capsid protein fused to a polyarginine tag cell-penetrating peptide on the C-terminus.
This part has been modified from the original (BBa_K2683012) so as to have the polyarginine tag replaced with a histidine tag, to allow for purification as a monomer for capsid assembly in vitro. It has also been codon optimized for E.coli.
The MS2 capsid protein comprises most of the capsid of the MS2 phage, as well as MS2 phage-like particles (PLPs). A single MS2 capsid requires 178 copies of the capsid protein, as one copy of maturation protein A (BBa_K3738000), which is critical in allowing the capsid to bind to the cell and deliver its cargo. The design of this part was based of part BBa_K2683012, which is an MS2 capsid protein fused to a polyarginine tag cell-penetrating peptide on the N-terminus. This part has been modified from the original (BBa_K2683012) so as to have a histidine tag placed on the C-terminus, to allow for purification as a monomer for capsid assembly in vitro. It has also been codon optimized for E.coli.
Lbu-Cas13a with an N-Terminal 6xHistidine Tag and C-Terminal Anionic Tag:
Cas13a is an enzyme originating from Leptotrichia buccalis (Lbu) which functions to cleave single-stranded RNAs (ssRNAs); particularly mRNAs. This function is achieved following protein-RNA complex formation with CRISPR RNA (crRNA) via crRNA backbone contacts with residues from the Helical-2, HEPN1, and Linker domains of Cas13a. The crRNA contains a spacer region coding for a direct repeat stem loop as well as a region complementary to target ssRNAs. Once the enzyme complex interacts with a target ssRNA, a structural conformation change occurs within the domains of the protein that permits active site formation for non-discriminate ssRNA cleavage (O’Connel et al., 2019).
Lbu-Cas13a with an N-terminal Anionic Tag and C-Terminal 6XHistidine Tag:
Cas13a is an enzyme originating from Leptotrichia buccalis (Lbu) which functions to cleave single-stranded RNAs (ssRNAs); particularly mRNAs. This function is achieved following protein-RNA complex formation with CRISPR RNA (crRNA) via crRNA backbone contacts with residues from the Helical-2, HEPN1, and Linker domains of Cas13a. The crRNA contains a spacer region coding for a direct repeat stem loop as well as a region complementary to target ssRNAs. Once the enzyme complex interacts with a target ssRNA, a structural conformation change occurs within the domains of the protein that permits active site formation for non-discriminate ssRNA cleavage (O’Connel et al., 2019).
Lbu-crRNA (Lbu signifying Leptotrichia buccalis, the source of the Cas13a protein and crRNA) is an RNA structure containing a direct-repeat stem loop, a recognition element for binding with the Lbu-Cas13a protein (see parts BBa_K3738020, BBa_K3738021, BBa_K3738023 and BBa_K3738024). The crRNA also contains a downstream complementary region, designed to base-pair with a target ssRNA sequence. Complex formation of crRNA-Cas13a occurs, and when the target sequence is perfectly paired with the crRNA, activation of the enzyme occurs and subsequent non-discriminate cleavage of collateral ssRNAs (O’Connell., 2019).
This crRNA was designed to bind with an important protein in the synthesis of harmful toxins called microcystins produced by blue-green algae (cyanobacteria) blooms. McyH is a gene in the Mcy gene cluster of Microcystic Aeruginosa and codes for a transporter protein. The protein is comprised of two homodimers each with a hydrophobic N-terminus domain and C-terminus containing an ATPase domain. Pearson et al., 2004 experimentally examined the impacts of impairing expression of this gene, and combined with bioinformatic data, found that McyH is likely a vital exporter of harmful microcystins and essential in their biosynthetic pathway. For this reason, we have designed crRNA to target the 5’-end of McyH in order to inhibit production of microcystins.
The P22 protein is originally from the P22 bacteriophage from Salmonella. The protein is able to properly form capsids (made of 420 subunits) when the capsid interacts with a scaffolding protein. With successful interaction, it forms capsids spontaneously. The P22 capsid protein has been previously used to encapsulate fluorescent proteins and CRISPR Cas9 systems.
MlrA, or microcystinase, cleaves the peptide backbone of MC-LR cyanotoxins produced by microcystis aeruginosa cyanobacteria. This crucial step results in the peptide losing the cyclic structure, and therefore the majority of its toxicity. Further degradation continues in an enzymatic pathway by interactions with other peptidolytic enzymes from the microcystinase gene cluster. This composite part includes the IPTG-inducible T7 promoter (BBa_J64997), RBS BBa_B0034, MlrA coding region BBa_K3738019, and double terminator BBa_B0015. The microcystinase coding region in this part has been codon-optimized for E.coli.
MlrA, or microcystinase, cleaves the peptide backbone of MC-LR cyanotoxins produced by Microcystis aeruginosa cyanobacteria. This crucial step results in the peptide losing the cyclic structure, and therefore the majority of its toxicity. Further degradation continues in an enzymatic pathway by interactions with other peptidolytic enzymes from the microcystinase gene cluster. This part originally comes from the Aalto-Helsinki 2016 iGEM team (Part BBa_K1907001), but was originally codon-optimized for yeast. We have modified this part by codon-optimizing it for use in E. coli. MlrA is known to be taken up into the periplasm with aid of transport protein MlrD (Saito et al., 2003), however for the mlrA sequence from Sphingomonas sp. USTB-05 the mlrD sequence is currently unavailable. Thus we have added an anionic tag for MS2-encapsulation.
This composite part includes the IPTG-inducible T7 promoter (BBa_J64997), RBS BBa_B0034, Lbu-Cas13a with C-Terminal Anionic Tag coding region (BBa_K3738020), and double terminator BBa_B0015.The part is codon-optimized for use in E. coli It is improved from the Lethbridge High School iGEM 2019's Parts BBa_K3001003, BBa_K3001000 and BBa_K3001002 by introducing the anionic MS2 phage-like-particle uptake anionic tag.
MS2 Bacteriophage Capsid Protein with Maturation Protein A:
The MS2 capsid protein comprises most of the capsid of the MS2 phage, as well as MS2 phage-like particles (PLPs). A single MS2 capsid requires 178 copies of the capsid protein, as one copy of maturation protein A (BBa_K3738000), which is critical in allowing the capsid to bind to the cell and deliver its cargo.
The first biobrick in this composite part includes the MS2 capsid protein fused to a polyarginine tag cell-penetrating peptide (BBa_K2683012) under the control of an strong arabinose-inducible promoter (BBa_K206000), as well as the RBS BBa_B0034 and double terminator BBa_B0015. The second biobrick contains maturation protein A (BBa_K3738000) under the control of weak constitutive promoter BBa_J23114, as well as the RBS BBa_B0034 and double terminator BBa_B0015. Given the high ratio of capsid protein to maturation protein A required, this design will result in higher production of the capsid protein, which can be induced once there is sufficient cell growth, resulting in self-assembly of functional MS2 PLPs.
Maturation Protein A is required in order for the capsid of the MS2 phage, and subsequently MS2-derived phage-like particles (PLP), to bind to bacterial pili. It is released from the capsid upon binding, and enters the cell along with the cargo to be delivered by the MS2 capsid. This composite part includes the IPTG-inducible T7 promoter (BBa_J64997), RBS BBa_B0034, maturation protein A coding region with N-Terminal histidine tag (BBa_K3738001), and double terminator BBa_B0015.
Maturation Protein A is required in order for the capsid of the MS2 phage, and subsequently MS2-derived phage-like particles (PLP), to bind to bacterial pili. It is released from the capsid upon binding, and enters the cell along with the cargo to be delivered by the MS2 capsid. This composite part includes the IPTG-inducible T7 promoter (BBa_J64997), RBS BBa_B0034, maturation protein A coding region with a C-terminal histidine tag (BBa_K3738002), and double terminator BBa_B0015. The maturation protein A coding region in this part has been codon-optimized for E.coli, and includes a histidine tag on the C-terminus for to allow for peptide purification.
Maturation Protein A is required in order for the capsid of the MS2 phage, and subsequently MS2-derived phage-like particles (PLP), to bind to bacterial pili. It is released from the capsid upon binding, and enters the cell along with the cargo to be delivered by the MS2 capsid. This composite part includes the IPTG-inducible T7 promoter (BBa_J64997), RBS BBa_B0034, maturation protein A coding region with a N-terminal histidine tag and TEV cleavage site (BBa_K3738003), and double terminator BBa_B0015. The maturation protein A coding region in this part has been codon-optimized for E.coli, and includes a histidine tag on the N-terminus for to allow for peptide purification. There is also a TEV cleavage site between the coding region and the histidine tag, in order to allow for histidine tag removal following purification if desired.
Maturation Protein A is required in order for the capsid of the MS2 phage, and subsequently MS2-derived phage-like particles (PLP), to bind to bacterial pili. It is released from the capsid upon binding, and enters the cell along with the cargo to be delivered by the MS2 capsid. This composite part includes the IPTG-inducible T7 promoter (BBa_J64997), RBS BBa_B0034, maturation protein A coding region with a C-terminal histidine tag and TEV cleavage site (BBa_K3738004), and double terminator BBa_B0015. The maturation protein A coding region includes a histidine tag on the C-terminus for to allow for peptide purification. There is also a TEV cleavage site between the coding region and the histidine tag, in order to allow for histidine tag removal following purification if desired.
The MS2 capsid protein comprises most of the capsid of the MS2 phage, as well as MS2 phage-like particles (PLPs). A single MS2 capsid requires 178 copies of the capsid protein, as one copy of maturation protein A (enter part number here), which is critical in allowing the capsid to bind to the cell and deliver its cargo. This composite part includes the IPTG-inducible T7 promoter (BBa_J64997), RBS BBa_B0034, MS2 capsid protein coding region (BBa_K3738005), and double terminator BBa_B0015. The MS2 capsid protein coding region in this part has been codon-optimized for E.coli, and includes a histidine tag on the N-terminus for to allow for peptide purification.
The MS2 capsid protein comprises most of the capsid of the MS2 phage, as well as MS2 phage-like particles (PLPs). A single MS2 capsid requires 178 copies of the capsid protein, as one copy of maturation protein A (BBa_K3738000), which is critical in allowing the capsid to bind to the cell and deliver its cargo. This composite part includes the IPTG-inducible T7 promoter (BBa_J64997), RBS BBa_B0034, MS2 capsid protein coding region with a C-terminal histidine tag (BBa_K3738006), and double terminator BBa_B0015. The MS2 capsid protein coding region in this part has been codon-optimized for E.coli, and includes a histidine tag on the C-terminus for to allow for peptide purification.
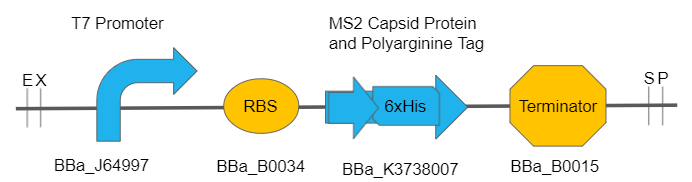
The MS2 capsid protein comprises most of the capsid of the MS2 phage, as well as MS2 phage-like particles (PLPs). A single MS2 capsid requires 178 copies of the capsid protein, as one copy of maturation protein A (BBa_K3738000), which is critical in allowing the capsid to bind to the cell and deliver its cargo. This composite part includes the IPTG-inducible T7 promoter (BBa_J64997), RBS BBa_B0034, MS2 capsid protein coding region fused to a polyarginine tag cell-penetrating peptide on the N-terminus and a histidine tag on the C-terminus (BBa_K3738007), and double terminator BBa_B0015. The MS2 capsid protein coding region in this part has been codon-optimized for E.coli, and includes a histidine tag on the C-terminus for to allow for peptide purification.
The MS2 capsid protein comprises most of the capsid of the MS2 phage, as well as MS2 phage-like particles (PLPs). A single MS2 capsid requires 178 copies of the capsid protein, as one copy of maturation protein A (BBa_K3738000), which is critical in allowing the capsid to bind to the cell and deliver its cargo. This composite part includes a coding region comprised of 2 copies of the MS2 capsid protein, joined by a histidine tag flanked by 2 linker sequences (BBa_K3738008). It also contains the IPTG-inducible T7 promoter (BBa_J64997), RBS BBa_B0034, and double terminator BBa_B0015. The MS2 capsid protein coding region in this part has been codon-optimized for E.coli.
The first biobrick in this composite part contains the coding region for the Lbu Cas13a enzyme, under control of the IPTG-inducible T7 promoter. It also includes multiple purification tags for Cas13a (BBa_K3001002), as well as the RBS BBa_B0034 and double terminator BBa_B0015. This part has already been submitted to the registry as the composite part BBa_K3001003.
This plasmid will also contain a biobrick with the sequence for the Cas13a crRNA, controlled by the same T7 promoter and followed by the terminator BBa_B0015. The crRNA coding region will contain a direct repeat stem loop sequence, as well as an short sequence that is found in the cyanobacteria genome. Once the Cas13a, containing the crRNA, recognizes the same sequence in the form of mRNA inside of cyanobacteria cells, it will become activated, and begin cleaving mRNA.
Lbu-Cas13a with an N-terminal 6xHistidine Tag and C-Terminal Anionic Tag:
This composite part includes the IPTG-inducible T7 promoter (BBa_J64997), RBS BBa_B0034, Lbu-Cas13a with an N-terminal 6xHistidine Tag and C-Terminal Anionic Tag coding region (BBa_K3738020), and double terminator BBa_B0015.The part is codon-optimized for use in E. coli It is improved from the Lethbridge High School iGEM 2019's Parts BBa_K3001003, BBa_K3001000 and BBa_K3001002 by introducing the anionic MS2 phage-like-particle uptake anionic tag as well as the transcriptional and translational regulators for optimal overexpression and 6XHistidine tag required for nickel affinity chromatography purification.
Lbu-Cas13a with an N-terminal Anionic Tag and C-Terminal Histidine Tag:
This composite part includes the IPTG-inducible T7 promoter (BBa_J64997), RBS BBa_B0034, Lbu-Cas13a with an N-terminal 6xHistidine Tag and C-Terminal Anionic Tag coding region (BBa_K3738020), and double terminator BBa_B0015.The part is codon-optimized for use in E. coli It is improved from the Lethbridge High School iGEM 2019's Parts BBa_K3001003, BBa_K3001000 and BBa_K3001002 by introducing the anionic MS2 phage-like-particle uptake anionic tag as well as the transcriptional and translational regulators for optimal overexpression and 6XHistidine tag required for nickel affinity chromatography purification.
The first biobrick in this composite part contains the coding region for the Lbu Cas13a enzyme, under control of the IPTG-inducible T7 promoter. It also includes multiple purification tags for Cas13a (BBa_K3001002), as well as the RBS BBa_B0034 and double terminator BBa_B0015. This part has already been submitted to the registry as the composite part BBa_K3001003.
This plasmid will also contain a biobrick with the sequence for the Cas13a crRNA, controlled by the same T7 promoter and followed by the terminator BBa_B0015. The crRNA coding region will contain a direct repeat stem loop sequence, as well as an short sequence that is found in the cyanobacteria genome. Once the Cas13a, containing the crRNA, recognizes the same sequence in the form of mRNA inside of cyanobacteria cells, it will become activated, and begin cleaving mRNA.
McyH complementary Lbu-crRNA:
This crRNA was designed to bind with an important protein in the synthesis of harmful toxins called microcystins produced by blue-green algae (cyanobacteria) blooms. McyH is a gene in the Mcy gene cluster of Microcystic Aeruginosa and codes for a transporter protein. The protein is comprised of two homodimers each with a hydrophobic N-terminus domain and C-terminus containing an ATPase domain. Pearson et al., 2004 experimentally examined the impacts of impairing expression of this gene, and combined with bioinformatic data, found that McyH is likely a vital exporter of harmful microcystins and essential in their biosynthetic pathway. For this reason, we have designed crRNA to target the 5’-end of McyH in order to inhibit production of microcystins.
The first biobrick in this composite part contains the coding region for the Lbu Cas13a enzyme, under control of the IPTG-inducible T7 promoter. It also includes multiple purification tags for Cas13a (BBa_K3001002), as well as the RBS BBa_B0034 and double terminator BBa_B0015. This part has already been submitted to the registry as the composite part BBa_K3001003.
This plasmid will also contain a biobrick with the sequence for the Cas13a crRNA, controlled by the same T7 promoter and followed by the terminator BBa_B0015. The crRNA coding region will contain a direct repeat stem loop sequence, as well as an short sequence that is found in the cyanobacteria genome. Once the Cas13a, containing the crRNA, recognizes the same sequence in the form of mRNA inside of cyanobacteria cells, it will become activated, and begin cleaving mRNA.
Lbu-Cas13a with an N-terminal 6xHistidine Tag and C-Terminal Anionic Tag and crRNA:
This crRNA was designed to bind with an important protein in the synthesis of harmful toxins called microcystins produced by blue-green algae (cyanobacteria) blooms. McyH is a gene in the Mcy gene cluster of Microcystic Aeruginosa and codes for a transporter protein. The protein is comprised of two homodimers, each with a hydrophobic N-terminus domain and C-terminus containing an ATPase domain. Pearson et al., 2004 experimentally examined the impacts of impairing expression of this gene, and combined with bioinformatic data, found that McyH is likely a vital exporter of harmful microcystins and essential in their biosynthetic pathway. For this reason, we have designed crRNA to target the 5’-end of McyH in order to inhibit production of microcystins.
This composite part includes the IPTG-inducible T7 promoter (BBa_J64997), RBS BBa_B0034, Lbu-Cas13a with an N-terminal 6xHistidine Tag and C-Terminal Anionic Tag coding region (BBa_K3738020), and double terminator BBa_B0015, followed by a second T7 promoter (BBa_J64997), McyH complementary Lbu-crRNA (BBa_K3738025), and a second BBa_B0015 terminator. The part is codon-optimized for use in E. coli It is improved from the Lethbridge High School iGEM 2019's Parts BBa_K3001003, BBa_K3001000 and BBa_K3001002 by introducing the anionic MS2 phage-like-particle uptake anionic tag as well as the transcriptional and translational regulators for optimal overexpression and 6XHistidine tag required for nickel affinity chromatography purification.
Lbu-Cas13a with an N-terminal Anionic Tag and C-Terminal 6xHistidine Tag and crRNA:
This crRNA was designed to bind with an important protein in the synthesis of harmful toxins called microcystins produced by blue-green algae (cyanobacteria) blooms. McyH is a gene in the Mcy gene cluster of Microcystic Aeruginosa and codes for a transporter protein. The protein is comprised of two homodimers, each with a hydrophobic N-terminus domain and C-terminus containing an ATPase domain. Pearson et al., 2004 experimentally examined the impacts of impairing expression of this gene, and combined with bioinformatic data, found that McyH is likely a vital exporter of harmful microcystins and essential in their biosynthetic pathway. For this reason, we have designed crRNA to target the 5’-end of McyH in order to inhibit production of microcystins.
This composite part includes the IPTG-inducible T7 promoter (BBa_J64997), RBS BBa_B0034, Lbu-Cas13a with an N-terminal Anionic Tag and C-Terminal 6xHistidine Tag coding region (BBa_K3738020), and double terminator BBa_B0015, followed by a second T7 promoter (BBa_J64997), McyH complementary Lbu-crRNA (BBa_K3738025), and a second BBa_B0015 terminator. The part is codon-optimized for use in E. coli It is improved from the Lethbridge High School iGEM 2019's Parts BBa_K3001003, BBa_K3001000 and BBa_K3001002 by introducing the anionic MS2 phage-like-particle uptake anionic tag as well as the transcriptional and translational regulators for optimal overexpression and 6XHistidine tag required for nickel affinity chromatography purification.